Rosalind Franklin and the Photograph That Changed Biology
On 6 May 1952, Rosalind Franklin and her graduate student Raymond Gosling produced the fifty-first X-ray diffraction photograph in their study of DNA at King's College London. It was not remarkable in how it was made — the technique was standard, the apparatus they used was one Franklin herself had refined and improved. What was remarkable was what it showed.
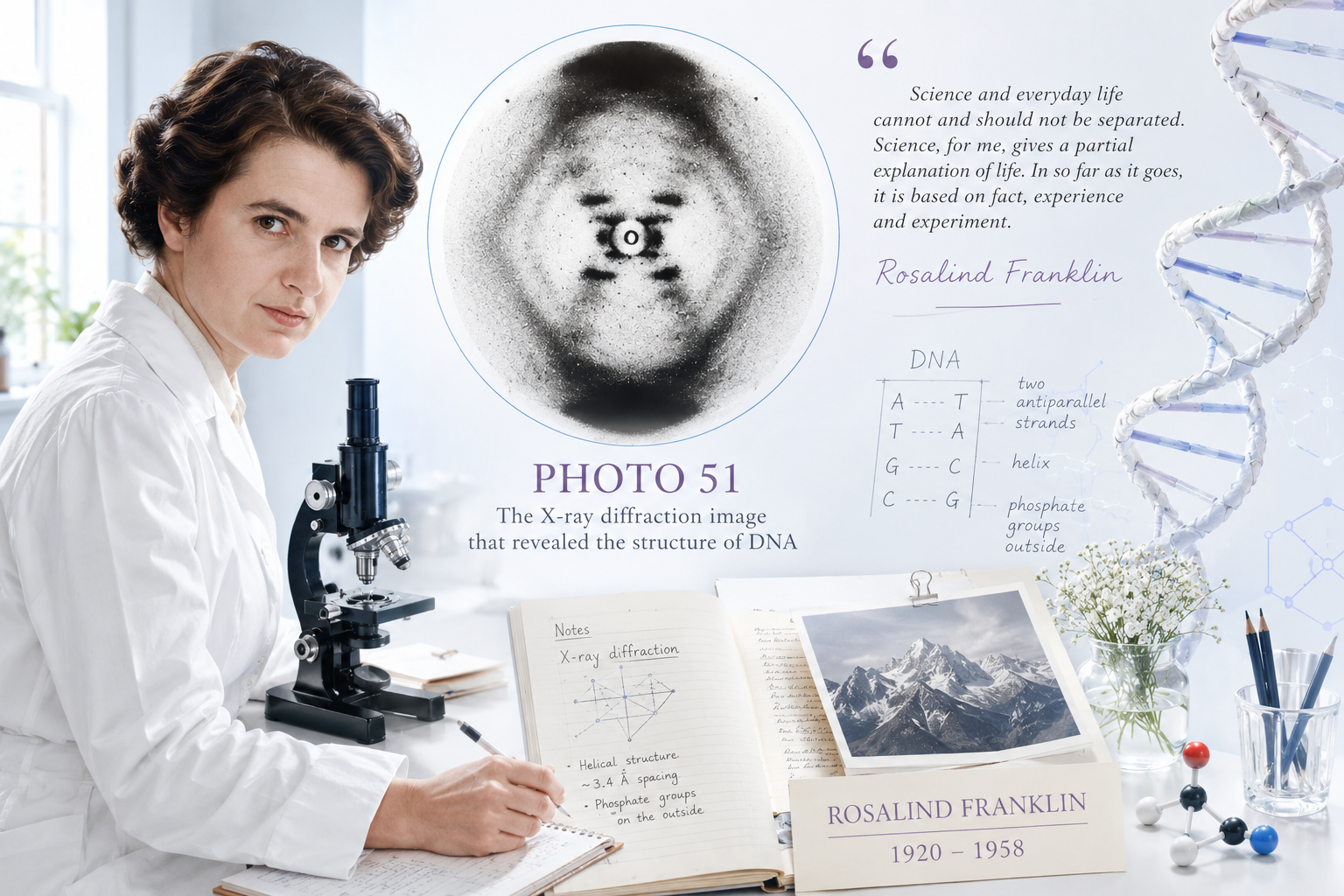
The photograph is blurry by the standards of modern imaging. It looks nothing like what most people expect a world-changing scientific document to look like. At its center is a dark X-shaped pattern, slightly asymmetric, surrounded by curved arcs of varying intensity. There are no labels, no annotations, no dramatic markings. Just the signature of a molecule written in scattered X-rays, captured on photographic film.
To anyone who understood X-ray crystallography, that image was a proof. It said, with the precision of physical law, that DNA was a helix — and it said it quantitatively, with the dimensions encoded directly in the geometry of the pattern. Understanding why requires following the physics into the photograph.
How X-Ray Crystallography Reads Structure
X-ray crystallography works on a deceptively simple principle: when radiation passes through a regular, repeating structure, it scatters — diffracts — in ways that encode the structure's geometry. The scattered rays interfere with each other, producing patterns of light and dark that carry precise mathematical information about the arrangement of atoms.
The governing relationship is the Bragg equation:
where is the wavelength of the X-rays, is the spacing between repeating planes of atoms in the molecule, is the angle at which strong scattering occurs, and is an integer. If you can measure the angles at which bright spots appear in the diffraction pattern, you can calculate the spacing — and from enough such measurements, reconstruct the three-dimensional geometry of the molecule.
For a helical molecule like DNA, the diffraction pattern takes a characteristic form. A helix can be mathematically decomposed as a series of repeating layers. Each layer contributes a set of reflections that appear along a specific row — called a layer line — in the diffraction pattern. The angle and spacing of these layer lines directly encode the pitch (the distance for one complete turn) and the radius of the helix. The pattern of intensities along each layer line encodes information about how the atoms are arranged within each turn.
The X-shape in Photo 51 is not decorative. It is a direct geometric consequence of helical structure. A helix scatters X-rays into a characteristic cross pattern because the repeating unit — one turn of the helix — projects onto a set of planes at angles that produce exactly that distribution of intensity. When Franklin saw the clean X in her photograph, she was seeing the mathematical fingerprint of helical geometry made visible by physics.
What Franklin Already Knew
By the time Photo 51 was taken, Franklin had already made a fundamental experimental insight that most accounts overlook.
Working with Gosling, she had discovered that DNA exists in two distinct forms depending on humidity. At lower water content — the A form — the diffraction pattern is complex and rich in detail, suitable for the kind of painstaking analysis that might yield detailed atomic positions. At higher water content — the B form, the conditions of living cells — the pattern is simpler and cleaner, with less information about individual atomic positions but with the helical structure unmistakably clear.
Photo 51 is of the B form. Franklin knew it was helical. From the layer line positions, she had already calculated the pitch of the helix: approximately 34 ångströms per complete turn. From the spacing of the reflections, she had inferred that there were approximately 10 base pairs per turn. From the distribution of intensity — particularly the strong signal toward the outer edges of the pattern — she had deduced that the phosphate groups lay on the outside of the helix, not the inside.
These were not guesses. They were the output of systematic mathematical analysis of diffraction data, performed with the care that characterizes genuinely rigorous experimental science. Franklin's notebooks from this period document the process in detail: measurements, calculations, comparisons with theory, the careful exclusion of alternative interpretations.
She had, in other words, already solved the major structural features of DNA — before Watson saw her photograph.
The Photograph Changes Hands
The circumstances under which Watson saw Photo 51 in January 1953 have been debated for decades, and the details matter more than the simplified version most people know.
Franklin herself had not shown Watson the photograph. Her colleague Maurice Wilkins — with whom she had a difficult and unresolved professional relationship, partly because neither had been clearly designated as leading the DNA project at King's — showed it to Watson during a meeting at King's College. Watson had not asked for it specifically, and Franklin did not know it had been shared.
Separately, Francis Crick saw a summary of Franklin's unpublished measurements in an MRC (Medical Research Council) progress report — a document circulated among scientific groups funded by the MRC, which included both King's and Cambridge. The report contained quantitative data from Franklin's analysis: the dimensions of the unit cell, the water content, the symmetry class. Crick, whose background was in X-ray crystallography, recognized immediately what these numbers implied.
Watson later described seeing Photo 51 as the moment of revelation: "The instant I saw the picture, my mouth fell open and my pulse began to race." His account in The Double Helix has shaped the popular narrative ever since. What it underemphasizes is that by the time he saw the photograph, Franklin had already extracted the most important information from it — and that the photograph's significance was legible to Watson precisely because Franklin's systematic analysis had established the framework for reading it.
Whether the sharing of the data was unethical, whether it constituted something that would today be called data misappropriation, whether Franklin was a collaborative or isolated figure in the complex politics of the King's College laboratory — these questions remain genuinely contested among historians of science. What is not contested is that Watson and Crick used Franklin's data without attribution, and did not acknowledge its role in their famous 1953 paper in Nature.
The Tobacco Mosaic Virus and What Came After
A dimension of Franklin's story that tends to disappear in the DNA narrative is what she did after leaving King's College in 1953.
At Birkbeck College, working with Aaron Klug, Franklin turned to viruses — specifically the tobacco mosaic virus and later the polio virus. The work was unhurried and methodologically meticulous, unclouded by the institutional tensions of King's College. It was, by most accounts of the scientists who knew her, the best and most rigorous research of her career.
She developed techniques for analyzing the structure of virus capsids using X-ray fiber diffraction, producing results of lasting importance. She traveled internationally to present her work. She was, in the years before her illness, a scientist in full possession of her powers and working in a field where her contribution was unambiguous.
She continued working through her illness. She died on 16 April 1958, at thirty-seven.
The Prize, the Rules, and What Cannot Be Corrected
In 1962, the Nobel Prize in Physiology or Medicine was awarded to Watson, Crick, and Wilkins for the discovery of the structure of DNA. Franklin had been dead for four years. Nobel Prizes are not awarded posthumously, and the rules have never permitted it — a policy that, in this case, means that a factual question about her eligibility has no answer.
Whether she would have been nominated, had she lived, is equally unanswerable. The Nobel committee's deliberations are sealed for fifty years, and the 1962 records are now available. They show that Franklin's name was not among the nominees that year — but the nominations for 1962 were submitted in 1961, three years after her death, and the rules at the time did not permit nominating deceased individuals.
What the historical record does show is that her contribution received essentially no acknowledgment in the 1953 papers that announced the double helix. It took Watson's 1968 memoir — a book so partial in its portrayal that Wilkins described it as "Jim's novel" — to bring her name back into public awareness, and in circumstances that were themselves deeply unfair to her.
Francis Crick, in later years, was unambiguous: Franklin's data had been critical. Without the quantitative measurements she had extracted from the diffraction patterns — the pitch, the number of bases per turn, the position of the phosphate groups — the double helix model could not have been built in the form it took.
What the Photograph Actually Proved
It is worth returning to the photograph itself, because the science matters as much as the story.
Photo 51 established that DNA is a helix with a pitch of 34 Å and approximately 10 base pairs per turn. These parameters constrained the model Watson and Crick built: the backbone spacing, the angles of the base pairs, the geometry of the proposed hydrogen bonds. The B-form pattern was also simpler than the A-form, which had previously resisted clean interpretation — Franklin's recognition that B-form DNA was the biologically relevant form and that it could be analyzed systematically was itself a scientific judgment, not merely a technical achievement.
The phosphate-outside conclusion was particularly important. Earlier model-building efforts, including Watson and Crick's failed attempt in 1951, had placed the phosphates on the inside — a position that made no chemical sense but that had not been definitively ruled out without the kind of quantitative crystallographic analysis Franklin was doing. Her data ruled it out. The correct model had to have the phosphates on the outside, the bases on the inside, the two strands running antiparallel — exactly the architecture of the double helix.
Franklin saw this. She had derived it from first principles, from the mathematics of diffraction, from careful experimental measurement. The photograph is the visible record of that derivation.
Precision as Scientific Virtue
Rosalind Franklin once wrote:
"Science and everyday life cannot and should not be separated. Science, for me, gives a partial explanation of life. In so far as it goes, it is based on fact, experience and experiment."
The sentence is characteristic — spare, precise, unsentimental. It describes a philosophy of science in which the obligation is to the evidence, not to a preferred outcome or a satisfying narrative. Franklin's careful insistence on distinguishing what the A-form data showed from what the B-form data showed, her reluctance to overstate conclusions that were not yet fully supported — these were not timidity or lack of insight. They were scientific integrity.
She saw farther than almost anyone around her, and she saw clearly, and she reported what she saw. That the field was not ready to credit the observation fully does not change what the observation was.
Photo 51 remains one of the most important scientific images ever produced — not because of the history surrounding it, though that history is significant, but because of what the image itself contains: a precise, quantitative, irreducible argument about the architecture of the molecule that carries the information of life. That argument was Rosalind Franklin's.
EisatoponAI
An independent intellectual publication exploring mathematics, AI, science, paradoxes, and the hidden structures behind reality.